Originally, cognitive performance improvement methods were focused on the senior population. Nootropics (cognitive enhancement pharmaceuticals or nutraceuticals) were being developed to aid victims of age-related decline, or neurodegenerative diseases.
In recent years, the focus of this fascinating field has broadened to include reducing stress-induced cognitive performance decline. This also widens the people who could benefit with people of all ages now seeking to enhance their cognitive performance.
In healthy individuals, stressors such as prolonged periods of demanding cognitive activity, physical activity, mental fatigue and workload stress, can trigger changes in the nervous system, affecting mental activities such as motivation and attention [1-3].
These stressors can lead to decline in performance and difficulty focusing on a task [4].
Many investigators have researched the efficacy of nootropics that claim to improve cognitive performance by modulating different aspects of the central nervous system [5].Only recently, has the focus shifted to the mechanism by which nootropics could reduce stress-induced cognitive impairment.
The molecular pathways involved in stress-induced cognitive impairment remain difficult to identify. To determine the optimal nutraceutical/nutritional approach, the biological mechanisms that facilitate improved cognition need to be understood.
If this pathway is discovered, stress induced cognitive impairment can be prevented naturally through nootropic supplementation.
.
Continuous aerobic exercise (the stress-induced model) has been positively associated with cognitive function and brain activity [6-9].
The physiological stress of endurance running has been linked with changes in brain-activity in the cortex region associated with impaired cognitive performance and mental fatigue [10-11].
We hypothesized that prolonged bouts of stress, such as aerobic exercise could activate pathways associated in stress-induced cognitive impairment. Endurance activity such as long-distance running would be a great model for unravelling factors involved in stress-induced cognitive decline.
In a recent study supported by the NRC-IRAP, OCE, and NSERC in collaboration with Carleton University, Staterra used a mechanistic stress-induced model (steady state physical exertion) to distinguish changes in blood amino acids levels. This innovative technique can detect amino acids patterns in response to stress.
Under normal conditions, blood concentration of individual amino acids remains at a relatively constant threshold level [12] . If amino acid fluctuations occur (such as during physical activity or mental fatigue) the body will attempt to correct this metabolic shift by breaking down muscle tissue since skeletal muscle is the only substantial option for storing amino acids [13-14].
It’s becoming very apparent that the plasma level of amino acids undergoes characteristic changes under various physiological conditions, and could, therefore, be a factor in stress-induced cognitive decline.
Amino acid analysis identified potential targets and more specifically a kinetic profile of amino acids during steady-state physical exertion linked to pathways related to stress. The kinetic profile of amino acids in this model are linked to various neuro-molecular pathways involved in cognition (Figure 1).

A unique study was performed to examine the amino acid kinetic patterns in healthy men and women recruited from the general population.
Thirty-four male and female participants between 18-45 years of age performed an 18 km run on a 400 m track, where dried blood spots (DBS) were collected before the run, at 6, 12, and 18 km, and 30 min post recovery.
Fasted blood samples were collected the morning of the trial and 24 hours after it. The DBS samples were analysed by Carleton University for several amino acids through mass-spectrometry.
The amino acid analysis identified potential targets, and more specifically, a kinetic profile of amino acids during steady-state physical exertion linked to pathways related to stress.
The kinetic profile of amino acids in this model are linked to various neuro-molecular pathways involved in cognition (Figure 1).
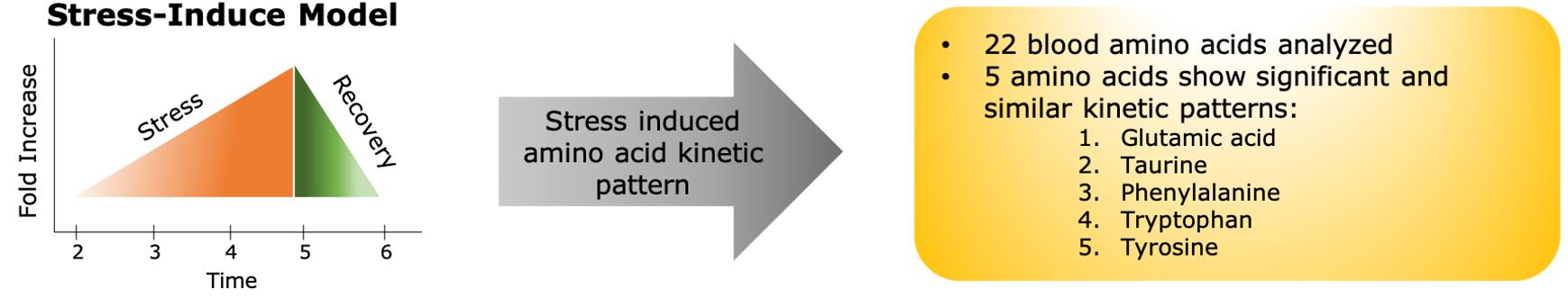
Figure 1: Stress-induced model. A novel mass spectrometry method developed to examine 22 amino acids extracted from dried blood samples of endurance athletes at set intervals over the course of a long-distance run shows a distinct pattern in 5 amino acids: glutamic acid, taurine, phenylalanine, tryptophan and tyrosine.
MindGain was developed from the findings of this study. Unlike the vast majority of supplements on the market, MindGain was developed through an understanding of the particular amino acids that are physiologically altered by stress.
The findings of this study show that stress triggers changes in amino acids and these changes have a specific kinetic profile that is statistically common amongst men and women and across the studied age range.
By analyzing the clusters of amino acids and their prominent role in neuro-molecular pathways, we identified significant kinetic patterns in neuro-molecular pathways related to cognitive functioning.
These pathways relate to:
- Neurotransmitter production
- Mitochondrial health
Repetitive multitasking can trigger a stress induced cognitive decline.
As humans, we naively believe we multitask effectively, however, research shows that our brains are not wired to rapidly switch between tasks. This skill cannot be developed even with dedicated training [15]
In fact, the more you multitask, the less you are able to perform well, concentrate, and learn.
Daily and repetitive attempts at multitasking can affect mood, cognitive performance and stress levels [16].
But MindGain has been proven to bridge this skills deficit.
During a recent market research study, MindGain was shown to improve multitasking efficacy in 29 healthy and active men and women with an average age of 34.
To determine the costs of mental ‘‘juggling’’, a task-switching paradigm popular with cognitive psychologists and neuroscientists was conducted [17].
Task-switching paradigms are routinely used to investigate aspects of cognitive control related to the updating, representation, and maintenance of frequently changing task rules.
The cued-task switching paradigm used in this market research is a common variant of the task-switching paradigm, in which the relevant task is signaled via an advance cue, thereby providing a measure of cognitive flexibility [17-18].
An example of this test is used in Dr. Delphie Dugal-Tessier’s TEDx talk titled “The Brain Hack to Get Your Life Back”.
By comparing how long it takes to switch between different tasks, a time cost ("task-switch cost”) can be calculated. Although task switch cost may be relatively small, it can add up to a large amount when people switch tasks repeatedly, thus causing mental blocks and decrease in productivity [19-21].
Those taking MindGain achieved a significant 50% decrease in the time it takes to switch from task to task. Task-switch cost is an indication of cognitive flexibility (Figure 2).

Figure 2:
MindGain decreases task-switch cost. Mean switching cost between baseline and 60 min after ingestion of MindGain,
Asterisk indicates significance (* p< 0.05) effect of MindGain on mean to baseline. Vertical capped lines atop bars indicates standard error of the mean.
Data analysed as a paired two-tailed sample t-test.
In addition, during the MindGain portion of the cued-task switch challenge, participants improved overall reaction time by an impressive 8% (Figure 3). The multitasking paradigm used in this study had an average accuracy rate of 91%, and with MindGain, participants were still able to improve their accuracy by 2% (Figure 4).
A single dose can improve cognitive flexibility ability across the board within 40 minutes of MindGain supplementation.

Vertical capped lines atop bars indicates standard error of the mean.

Figure 4:
MindGain improves accuracy. Percentage of correct responses during multitag task following ingestion of MindGain indicates significance (* p< 0.05) effect of MindGain on mean to baseline. Vertical capped lines atop bars indicates standard error of the mean
Data analysed as a paired two-tailed sample t-test.
These results harmonize with the literature on amino acids involved in the synthesis of neurotransmitters.
For example, it’s broadly studied and accepted that the catecholamine system, and more specifically dopamine, influences cognitive flexibility in healthy young adults as well as a healthy aging population [22-27].
Supplementation with the amino acid tyrosine, the precursor of dopamine, has been shown to improve performance in challenging tasks related to the catecholamine pathway such as multitasking and working memory [28-39].
The MindGain market research agrees with the scientific literature and strongly indicates that certain amino acids, like tyrosine, play a key role in cognitive performance.
MindGain goes one step further to incorporate the nutritional constituents needed to mitigate stress response, enhance focus and cognitive performance.
All of MindGain’s benefits are listed below. Clicking on each one will reveal a list of supporting studies.
1. de Lange FP, Faber LG, Maurits NM, Lorist MM. Mental fatigue affects visual selective attention. PloS ONE (2012) 7:e48073.
2. Xu R., et al. How Physical Activities Affect Mental Fatigue Based on EEG Energy, Connectivity, and Complexity. Front Neurol. 2018; 9: 915.
3. Fan J & Smith AP. The Impact of Workload and Fatigue on Performance. Human Mental Workload: Model and applications. (2017) pp 90-105.
4. Boksem MA, Meijman TF, Lorist MM. Mental fatigue, motivation and action monitoring. Biol Psychol. (2006) 72:123–32.
5. Suliman NA et al. Establishing Natural Nootropics: Recent Molecular Enhancement Influenced by Natural Nootropic. id Based Complement Alternat Med 2016; 2016:4391375
6. Holzschneider K, Wolbers T, Roder B & Hotting K. Cardiovascular fitness modulates brain activation associated with spatial learning. Neuroimage. 2011;59:3003-3014
7. Erickson KI, Weinstein AM, Sutton BP. Beyond vascularization: aerobic fitness is associated with N-acetylaspartate and working memory. Brain Behav. 2012;2:32-41.
8. Head D, Singh T, Bugg JM. The moderating role of exercise on stress-related effects on the hippocampus and memory in later adulthood. Neuropsychology. 2012;26:133-143.
9. Zoeller RF. Exercise and cognitive function: can working out train the brain, too? Am J Lifestyle Med. 2010;4:397-409.
10. Schiphof-Godart L, Roelands B & Hettinga FJ. Drive in Sports: How Mental Fatigue Affects Endurance Performance. Front Psychol. 2019;9:1383.
11. Martin K., Meeusen R., Thompson K. G., Keegan R., Rattray B. (2018). Mental fatigue impairs endurance performance: a physiological explanation. Sports Med. 48 2041–2051.
12. Ra, S.G., et al., Combined effect of branched-chain amino acids and taurine supplementation on delayed onset muscle soreness and muscle damage in high-intensity eccentric exercise. J Int Soc Sports Nutr, 2013. 10(1): p. 51
13. Moore, D.R., Keeping older muscle "young" through dietary protein and physical activity. Adv Nutr, 2014. 5(5): p. 599S-607S.
14. Stoet, G. & Snyder, LH. Extensive practice does not eliminate human switch costs. Cogn Affect Behav Neurosci. (2007) Sep;7(3):192-7.
15. Rosen, L. Cheeber, NA & Carrier, LM. The Wiley Handbook of Psychology, Technology, and Society. (2015).
John Wile & Sons, Ltd.
16. Mayr U. (2006). What matters in the cued task-switching paradigm: tasks or cues? Psychon Bull Rev. 2006 Oct;13(5):794-9.
17. Meiran, N. (1996). Reconfiguration of processing mode prior to task performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 22(6), 1423–1442.
18. American Psychological Association (https://www.apa.org/research/action/multitask)
19. Meyer, D. E. & Kieras, D. E. (1997a). A computational theory of executive cognitive processes and multiple-task performance: Part 1. Basic mechanisms. Psychological Review, 104, 3-65.
20. Meyer, D. E. & Kieras, D. E. (1997b). A computational theory of executive cognitive processes and multiple-task performance: Part 2. Accounts of psychological refractory-period phenomena. Psychological Review, 104, 749-791.
21. P.S. Goldman-Rakic, E.C. Muly, G.V.Williams. D1 receptors in prefrontal cells and circuits Brain Res. Rev., 31 (2000), pp. 295-301
22. E.C. Muly III, K. Szigeti, P.S.Goldman-Rakic. D1 receptor in interneurons of macaque prefrontal cortex: distribution and subcellular localization J. Neurosci., 18 (1998), pp. 10553-10565
23. Jonkees BJ, Hommel B, Kuhn S, Colzato LS. Effect of tyrosine supplementation on clinical and healthy populations under stress or cognitive demands--A review. J Psychiatr Res. 2015 Nov;70:50-7.
24. Banderet LE, Lieberman HR. Treatment with tyrosine, a neurotransmitter precursor, reduces environmental stress in humans. Brain Res Bull, 1989; 22(4):759–62,
25. Lieberman HR. Nutrition, brain function and cognitive performance. Appetite 2003; 40(3):245-54
26. Neri DF, Wiegmann D, Stanny RR, Shappell SA, McCardie A, McKay DL The effects of tyrosine on cognitive performance during extended wakefulness. Aviat Space Environ Med 1995; 66(4):313–9.
27. Magill RA, Waters WF, Bray GA, et al. Effects of tyrosine, phentermine, caffeine D-amphetamine, and placebo on cognitive and motor performance deficits during sleep deprivation. Nutr Neurosci 2003; 6(4): 237–46.
28. Deijen JB, Wientjes CJ, Vullinghs HF, Cloin PA, Langefeld JJ Tyrosine improves cognitive performance and reduces blood pressure in cadets after one week of a combat training course Brain Res Bull 1999; 48(2):203–9.
29. Attipoe S et al. Tyrosine for Mitigating Stress and Enhancing Performance in Healthy Adult Humans, a Rapid Evidence Assessment of the Literature. Military Medicine 2015; 180(7): 754-765.
30. D. Shurtleff, J.R. Thomas, J. Schrot, K.Kowalski, R. Harford Tyrosine reverses a cold-induced working memory deficit in humans Pharmacol. Biochem. Be, 47 (1994), pp. 935-941.
31. C.R. Mahoney, J. Castellani, F.M.Kramer, A. Young, H.R. Lieberman Tyrosine supplementation mitigates working memory decrements during cold exposure Physiol. Behav., 92 (2007), pp. 575-582.
32. J.R. Thomas, P.A. Lockwood, A. Sing, P.A. Deuster Tyrosine improves working memory in a multitasking environment Pharmacol. Biochem. Behav., 64(1999), pp. 495-500
33. L.S. Colzato, B. Jongkees, R. Sellaro, B. Hommel Working memory reloaded: tyrosine repletes updating in the N-Back task Front. Behav. Neurosci., 7 (2013), p. 200,
34. Berry AS, et al. Aging affects dopaminergic neural mechanisms of cognitive flexibility. 2016. J Neurosci. 36(50):12559-1269
35. Wang Y, Chan GL, Holden JE, Dobko T, Mak E, Schulzer M, Huser JM, Snow BJ, Ruth TJ, Calne DB, Stoessl AJ (1998) Age-dependent decline of do- pamine D1 receptors in human brain: a PET study. Synapse 30:56–61.
36. Erixon-Lindroth N, Farde L, Wahlin TB, Sovago J, Halldin C, Backman L (2005) The role of the striatal dopamine transporter in cognitive aging. Psychiatry Res 138:1–12.
37. Samanez-Larkin GR, Buckholtz JW, Cowan RL, Woodward ND, Li R, Ansari MS, Arrington CM, Baldwin RM, Smith CE, Treadway MT, Kessler RM, Zald DH (2013) A thalamocorticostriatal dopamine network for psychostimulant-enhanced human cognitive flexibility. Biol Psychiatry 74:99 –105.
38. Stelzel C, Basten U, Montag C, Reuter M, Fiebach CJ (2010) Frontostriatal involvement in task switching depends on genetic differences in d2 recep- tor density. J Neurosci 30:14205–14212.
39. Jin G et al,. Changes in plasma and tissue amino acid levels in an animal model of complex fatigue. Nutrition. 2009 May;25(5):597-607.